
“Picture yourself hiking through the woods or walking across a lawn,” says Elizabeth Haswell, PhD, assistant professor of biology in Arts & Sciences at Washington University in St. Louis. “Now ask yourself: Do the bushes know that someone is brushing past them? Does the grass know that it is being crushed underfoot? Of course, plants don’t think thoughts, but they do respond to being touched in a number of ways.”
“It’s clear,” Haswell says, “that plants can respond to physical stimuli, such as gravity or touch. Roots grow down, a ‘sensitive plant’ folds its leaves, and a vine twines around a trellis. But we’re just beginning to find out how they do it,” she says.
In the 1980s, work with bacterial cells showed that they have mechanosensitive channels, tiny pores in the cell’s membrane that open when the cell bloats with water and the membrane is stretched, letting charged atoms and other molecules rush out of the cell. Water follows the ions, the cell contracts, the membrane relaxes, and the pores close.
Genes encoding seven such channels have been found in the bacterium Escherichia coli and 10 in Arabidopsis thaliana, a small flowering plant related to mustard and cabbage. Both E. coli and Arabidopsis serve as model organisms in Haswell’s lab.
She suspects that there are many more channels yet to be discovered and that they will prove to have a wide variety of functions.
Recently, Haswell and colleagues at the California Institute of Technology, who are co-principal investigators on an National Institutes of Health (NIH) grant to analyze mechanosensitive channels, wrote a review article in order to “get their thoughts together” as they prepared to write the grant renewal. The review appeared in the Oct. 11 issue of Structure.
Swelling bacteria might seem unrelated to folding leaflets, but Haswell is willing to bet they’re all related and that mechanosensitive ion channels are at the bottom of them all. After all, plant movements — both fast and slow — are ultimately all hydraulically powered; where ions go the water will follow.
Giant E. coli cells
The big problem with studying ion channels has always been their small size, which poses formidable technical challenges.
Early work in the field, done to understand the ion channels whose coordinated opening and closing creates a nerve impulse, was done in exceptionally large cells: the giant nerve cells of the European squid, which had projections big enough to be seen with the unaided eye.
Experiments with these channels eventually led to the development of a sensitive electrical recording technique known as the patch clamp that allowed researchers to examine the properties of a single ion channel. Patch clamp recording uses as an electrode a glass micropipette that has an open tip. The tip is small enough that it encloses a “patch” of cell membrane that often contains just one or a few ion channels.
Patch clamp work showed that there were many different types of ion channels and that they were involved not just in the transmission of nerve impulses but also with many other biological processes that involve rapid changes in cells.
Mechanosensitive channels were discovered when scientists started looking for ion channels in bacteria, which wasn’t until the 1980s because ion channels were associated with nerves and bacteria weren’t thought to have a nervous system.
In E. coli, the ion channels are embedded in the plasma membrane, which is inside a cell wall, but even if the wall could be stripped away, the cells are far too small to be individually patched. So the work is done with specially prepared giant bacterial cells called spherophlasts.
These are made by culturing E. coli in a broth containing an antibiotic that prevents daughter cells from separating completely when a cell divides. As the cells multiply, “snakes” of many cells that share a single plasma membrane form in the culture. “If you then digest away the cell wall, they swell up to form a large sphere,” Haswell says.
Not that spheroplasts are that big. “We’re doing most of our studies in Xenopus oocytes (frog eggs), whose diameters are 150 times bigger than those of spheroplasts,” she says.
Three mechanosensitive channel activities
To find ion channels in bacteria, scientists did electrophysiological surveys of spheroplasts. They stuck a pipette onto the spheroplast and applied suction to the membrane as they looked for tiny currents flowing across the membrane.
“What they found was really amazing,” Haswell says. “There were three different activities that are gated (triggered to open) only by deformation of the membrane.” (They were called “activities” because nobody knew their molecular or genetic basis yet.)
The three activities were named mechanosensitive channels of large (MscL), small (MscS) and mini (MscM) conductance. They were distinguished from one another by how much tension you had to introduce in order to get them to open and by their conductance.
One of the labs working with spheroplasts was led by Ching Kung, PhD, at the University of Wisconsin-Madison. The MscL protein was identified and its gene was cloned in 1994 by Sergei Sukharev, PhD, then a member of Kung’s lab. His tour-de-force experiment, Haswell says, involved reconstituting fractions of the bacterial plasma membrane into synthetic membranes (liposomes) to see whether they would confer large-channel conductance.
In 1999, the gene encoding MscS was identified in the lab of Ian Booth, PhD, at the University of Aberdeen. Comparatively, little work has been done on the mini channel, which is finicky and often doesn’t show up, Haswell says, though a protein contributing to MscM activity was recently identified by Booth’s group.
Once both genes were known, researchers did knockout experiments to see what happened to bacteria that didn’t have the genes needed to make the channels. What they found, says Haswell, was that if both the MscL and MscS genes were missing, the cells could not survive “osmotic downshock,” the bacterial equivalent of water torture.
“The standard assay,” Haswell says, “is to grow the bacteria for a couple of generations in a very salty broth, so that they have a chance to balance their internal osmolyte concentration with the external one.” (Osmolytes are molecules that affect osmosis, or the movement of water into and out of the cell.) “They do this,” she says, “by taking up osmolytes from the environment and by making their own.”
“Then,” she says, “you take these bacteria that are chockfull of osmolytes and throw them into fresh water. If they don’t have the MscS and MscL proteins that allow them to dump ions to avoid the uncontrolled influx of water, they don’t survive.” It’s a bit like dumping saltwater fish into a freshwater aquarium.
Why are there three mechanosenstivie channel activities? The currently accepted model, Haswell says is that the channels with the smaller conductances are the first line of defense. They open early in response to osmotic shock so that the channel of large conductance, through which molecules the cell needs can escape, doesn’t open unless it is absolutely necessary. The graduated response thus gives the cell its best chance for survival.
Crystallizing the proteins
The next step in this scientific odyssey, figuring out the proteins’ structures, also was very difficult. Protein structures are traditionally discovered by purifying a protein, crystallizing it out of a water solution, and then bombarding the crystal with X-rays. The positions of the atoms in the protein can be deduced from the X-ray diffraction pattern.
In a sense crystallizing a protein isn’t all that different from growing rock candy from a sugar solution, but, as always, the devil is in the details. Protein crystals are much harder to grow than sugar crystals and, once grown, they are extremely fragile. They even can even be damaged by the X-ray probes used to examine them.
And to make things worse MscL and MscS span the plasma membrane, which means that their ends, which are exposed to the periplasm outside the cell and the cytoplasm inside the cell, are water-loving and their middle sections, which are stuck in the greasy membrane, are repelled by water. Because of this double nature it is impossible to precipitate membrane proteins from water solutions.
Instead the technique is to surround the protein with what have been characterized as “highly contrived detergents,” that protect them — but just barely — from the water. Finding the magical balance can take as long as a scientific career.
The first mechanosensitive channel to be crystallized was MscL — not the protein in E. coli but the analogous molecule (a homolog) from the bacterium that causes tuberculosis. This work was done in the lab of one of Haswell’s co-authors, Douglas C. Rees a Howard Hughes investigator at the California Institute of Technology.The third co-author, Rob Phillips, also of CalTech, earned his PhD in physics at Washington University in St. Louis in 1989.
MscS from E. coli was crystallized in the Rees laboratory several years later, in 2002, and an MscS protein with a mutation that left it stuck in the presumed open state was crystallized in the Booth laboratory in 2008. “So now we have two crystal structures for MscS and two (from different bacterial strains) for MscL,” Haswell says.
Of plants and mutants

Up to this point, mechanosensitive channels might not seem all that interesting because they seemed to be restricted to bacteria.
However, says, Haswell, in the early 2000s, scientists began to compare the genes for the bacterial channels to the genomes of other organisms and they discovered that there are homologous sequences not just in other bacteria but also in some multicellular organisms, including plants.
“This is where I got involved,” she says. “I was interested in gravity and touch response in plants. I saw these papers and thought these homologs were great candidates for proteins that might mediate those responses.”
“There are 10 MscS-homologs in Arabidopsis and no MscL homologs,” she says. “What’s more, different homologs are found not just in the cell membrane but also in chloroplast and mitochondrial membranes. “
The chloroplast is the light-capturing organelle in a plant cell and the mitochondria is its power station; both are thought to be once-independent organisms that were engulfed and enslaved by cells which found them useful. Their membranes are vestiges of their free-living past.
The number of homologs and their locations in plant cells suggests these channels do much more than prevent the cells from taking on board too much water.

So what exactly were they doing? To find out Haswell got online and ordered Arabidopsis seeds from the Salk collection in La Jolla, Calif., each of which had a mutation in one of the 10 channel genes.
From these mutants she’s learned that two of the ten channels control chloroplast size and proper division as well as leaf shape. Plants with mutations in these two MscS channel homologs have giant chloroplasts that haven’t divided properly. The monster chloroplasts garnered her lab the cover of the August issue of The Plant Cell.
“We showed that bacteria lacking MscS and MscL don’t divide properly either,”Haswell says, “so the link between these channels and division is evolutionarily conserved.”
The big idea
But Haswell and her co-authors think they are only scratching the surface. “We are basing our understanding of this class of channels on MscS itself, which is a very reduced form of the channel,” she says. “It’s relatively tiny.”
“But we know that some of the members of this family have long extensions that stick out from the membrane either outside or inside the cell. We suspect this means that the channels not only discharge ions, but that they also signal to the whole cell in other ways. They may be integrated into common signaling pathways, such as the cellular osmotic stress response pathway.
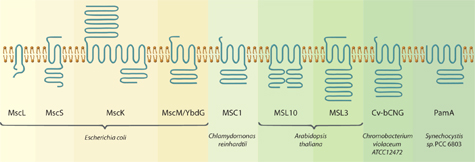
E. coli’s MscL and MscS channels (left), says Haswell, may be reduced forms of mechanosensitive channels. Many of the other known channels have extensions either outside or inside the cell that suggest they are up to something more complex than MscL and MscS. (The small brown clothespins between blue squiggles are a schematic representation of the plasma membrane the channels bridge.)
We think we may be missing a lot of complexity by focusing too exclusively on the first members of this family of proteins to be found and characterized,” she says. “We think there’s a common channel core that makes these proteins respond to membrane tension but that all kinds of functionally relevant regulation may be layered on top of that.”
“For example,” she says, “there’s a channel in E. coli that’s closely related to MscS that has a huge extension outside the cell that makes it sensitive to potassium. So it’s a mechanosensitive channel but it only gates in the presence of potassium. What that’s important for, we don’t yet know, but it tells us there are other functions out there we haven’t studied.”
What about the sensitive plant?
So are these channels at the bottom of the really fast plant movements like the sensitive plant’s famous touch shyness? (To see a movie of this and other “nastic” (fast) movements, go to the Plants in Motion site maintained by Haswell’s colleague Roger P. Hangartner of Indiana University).
Haswell is circumspect. “It’s possible,” she says. “In the case of Mimosa pudica there’s probably an electrical impulse that triggers a loss of water and turgor in cells at the base of each leaflet, so these channel proteins are great candidates.